Resolving the Display–Storage Contradiction Through the Gyrus
BY: OMOLAJA MAKINEE
One of the deepest architectural contradictions in modern Neuroscience emerged not from a lack of data, but from a failure of classification.
For over a century, Neuroscience accumulated extraordinary knowledge regarding memory, emotion, perception, learning, behavioural reinforcement, attention, and consciousness. Yet despite this expanding mountain of information, a fundamental problem remained unresolved. Structures performing radically different operations continued to be grouped inside the same anatomical civilisation.
The result was a conceptual compression that silently destabilised Behavioural science.
Memory indexing systems sat beside symbolic display systems. Emotional weighting systems sat beside conscious awareness systems. Behavioural packaging systems sat beside sensory rendering systems. Recording machinery sat beside projection machinery. Everything was placed inside the broad territorial umbrella of the traditional Telencephalon.
Psychextrics identifies this as the Display–Storage Contradiction.
The contradiction emerged because Neuroscience fused together two fundamentally different classes of biological hardware while treating them as members of the same operational family.
The key to resolving this contradiction lies in one overlooked anatomical feature: the gyrus.
1. The Forgotten Significance of the Gyrus
A gyrus is one of the raised ridges visible across the folded surface of the brain.
Traditionally, gyri have been organised according to their physical location within the major cerebral lobes. Frontal gyri belonged to the frontal lobe. Temporal gyri belonged to the temporal lobe. Parietal gyri belonged to the parietal lobe.
While useful for anatomical navigation, this method says very little about behavioural architecture.
Psychextrics approaches the problem differently. Instead of asking where a gyrus is located geographically, it asks a more important question:
What operational role does the gyrus perform?
When viewed through this lens, a remarkable pattern emerges. Every major gyrus belongs to one of only two broad functional territories. Either it participates in behavioural integration and signal compilation. Or it participates in behavioural display and symbolic rendering. There is no middle ground.
The moment this distinction is recognised, the Display–Storage Contradiction begins to dissolve.
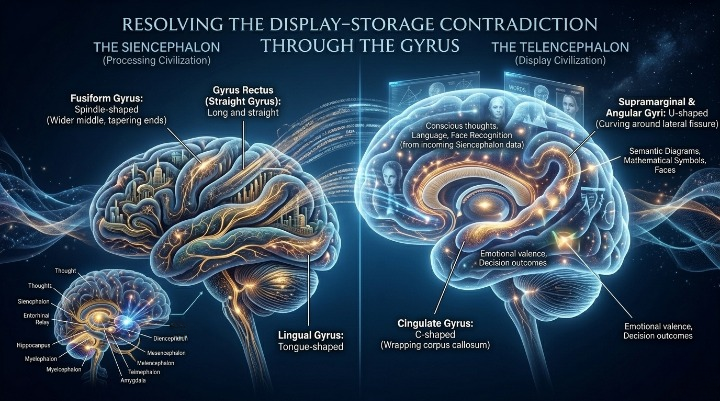
2. Two Civilisations Occupying One Surface
Within the 6-Cephalon Model, gyri exist exclusively within two cephalic territories. The first territory is the Siencephalon. The second territory is the Telencephalon.
Although both contain folded cortical structures, their functions are fundamentally different. The Siencephalon operates as a processing civilisation. The Telencephalon operates as a display civilisation.
The Siencephalic gyri function as backend engineering systems. Their purpose is to receive incoming behavioural streams from the lower cephalons, organise those streams, assign significance to them, stabilise them, and prepare them for conscious presentation.
The Telencephalic gyri perform none of these operations. Instead, they act as rendering surfaces. Their purpose is to convert already-packaged behavioural information into conscious experience.
- One compiles. The other displays.
- One constructs. The other reveals.
- One prepares behavioural reality. The other projects behavioural reality.
This distinction establishes an operational boundary that remains absolute regardless of how strongly the two systems communicate.
3. The Architecture of the Processing Engine
The Siencephalic gyri form a hidden ring of behavioural translators. Each serves as a specialised relay system for lower cephalic infrastructures.
- The Cingulate Gyrus functions as a visceral conflict and metabolic valuation relay.
- The Parahippocampal Gyrus serves as a spatial and environmental translation gateway.
- The Dentate Gyrus performs pattern separation and memory indexing operations.
- The Subcallosal Gyrus links visceral survival states to chemical and olfactory behavioural streams.
These structures do not generate conscious imagery. They do not produce internal narration. They do not create symbolic awareness. Instead, they operate as behavioural compilers. They transform raw cephalic traffic into organised behavioural packets suitable for later display.
Their role resembles that of a film production studio. They create the content. They prepare the sequence. They organise the material. But they do not project the movie.
4. The Architecture of the Display Screen
The Telencephalic gyri perform the opposite task.
- The Precentral Gyrus renders physical action commands.
- The Postcentral Gyrus renders tactile awareness.
- The Superior Temporal Gyrus renders auditory experience.
- The Angular Gyrus renders symbolic matching.
- The Fusiform Gyrus renders facial recognition.
- The Orbitofrontal Gyri render conscious valuation.
- The Insular Gyri render interoceptive awareness.
These structures do not generate the behavioural packets they display. They simply translate incoming siencephalic compilations into conscious form.
Like the pixels on a television screen, they illuminate information that has already been prepared elsewhere. The screen is essential. But it is not the source of the broadcast.
5. The Great Network Mistake
The confusion persisted because twentieth-century Neuroscience increasingly abandoned structural hierarchy in favour of connectivity.
As neuroimaging technologies advanced, researchers became fascinated with networks. Structures that activated simultaneously were grouped together. Regions connected by dense white matter pathways were classified as functional partners.
Synchronisation became the dominant explanatory principle. This produced an epistemological substitution.
The question shifted from:
“What is this structure built to do?”
To:
“What other structures does it communicate with?”
The distinction appears subtle. Its consequences were enormous.
When the Anterior Cingulate and the Prefrontal Cortex activated together during pain, they were placed inside the same network.
When the Posterior Cingulate synchronised with Medial Prefrontal regions, they became members of the same default mode system.
When Parahippocampal structures communicated extensively with Temporal regions, they were merged into shared cognitive frameworks.
The communication became mistaken for equality. The connection became mistaken for identity. The telephone line became mistaken for the office itself.
6. Why Synchronisation Cannot Erase Architecture
Psychextrics argues that synchronisation does not abolish hierarchy. Two structures can fire simultaneously while performing entirely different jobs.
A conductor and a violinist may act in perfect synchrony during a symphony. Yet one is coordinating the performance while the other is producing sound.
A database server and a computer monitor may exchange information continuously. Yet one stores data while the other displays it.
The intensity of communication does not erase the distinction between storage and display. The same principle applies to the gyri.
It does not matter how intensely a Siencephalic gyrus and a Telencephalic gyrus synchronise their firing rates. They remain separated by an unbridgeable operational boundary.
One processes behavioural reality. The other displays behavioural reality. One functions as backend machinery. The other functions as frontend projection.
No degree of synchronisation transforms a compiler into a screen. No degree of communication transforms a screen into a compiler.
7. Restoring Behavioural Sequence
Once this boundary is restored, Behavioural science regains something it has lacked for decades:
Sequence.
Behaviour can once again be tracked through a coherent architectural pathway.
Signals originate within the lower cephalons. Those signals enter specialised Siencephalic gyri for indexing, valuation, separation, stabilisation, and compilation. The finished behavioural package is then transmitted upward. Only at the final stage does the Telencephalic mantle illuminate the result as conscious awareness.
The order becomes visible. The hierarchy becomes visible. The assembly process becomes visible.
The brain ceases to look like a flat political network and begins to resemble what it actually is:
A layered biological civilisation operating through specialised divisions of labour.
Conclusion: Connectivity Is Not Architecture
The Display–Storage Contradiction persisted because Behavioural science increasingly prioritised connectivity over structure.
- As networks became fashionable, architectural boundaries weakened.
- As synchronisation became central, hierarchy disappeared.
- As communication maps expanded, behavioural sequence collapsed.
Psychextrics resolves this contradiction by restoring strict operational demarcation between the gyri of the Siencephalon and the gyri of the Telencephalon.
The Siencephalic gyri compile behavioural reality. The Telencephalic gyri display behavioural reality.
One is the engine room. The other is the projection screen.
One writes the behavioural ledger. The other makes that ledger visible.
And no amount of synchronised firing can erase the fundamental distinction between the place where reality is assembled and the place where reality is shown.
Back to: 👇